




果蝇大脑的Brainbow图像(图片来源:HHMI)
多久以来,科学家对记忆的探究都避开了“遗忘”的话题:人们认为“遗忘”就是记忆的消失,“防止遗忘”就是增强记忆的最好方法。但随着近几年神经科学界对遗忘机制的研究,一片复杂的图景逐渐浮现:遗忘并非简单的“消除记忆”。“话在嘴边”(tip-of-the-tongue)现象就是一个例子:突然忘记的名字、生日、手机号,会在一段时间过后突然出现在我们的脑海里,而最近刊登于《自然》的一项研究,终于开始探索这种现象的神经学本质。
德彪西说过,“是音符间的间隙造就了音乐。”(Music is the space between the notes.)这句话放在记忆上也毫不为过:是片段之间的空白造就了记忆——没有遗忘,记忆便无从谈起。
遗忘(forgetting)是生物记忆的必经之路。在2019年的一篇《自然》的评论文章中,科普作家劳伦·格拉维茨(Lauren Gravity)写道:“虽然长久以来,遗忘都被视作记忆失灵的例子,但研究人员们已经逐渐意识到,遗忘对大脑的正常运作至关重要。”格拉维茨的观点在神经科学界找到了共鸣。近几年,无数的遗忘研究涌现。这些研究不仅说明了遗忘是大脑主动发起的生物过程,还为利用遗忘治疗诸如创伤后压力症候群(PTSD)等精神疾病提供了新的思路。
然而,遗忘不只有一种形式。除了长期的、内源性的遗忘(intrinsic forgetting),生物还会经历短期的、瞬时性的遗忘(transient forgetting)。这样的现象在生活中并不少见。例如在一些情况下,当别人问起你的生日时,你会突然“卡壳”,好久也想不起来。同样的情况也可能出现在对熟人名字的回忆上,当然最糟糕的,还要数在考试时出现瞬时遗忘。有趣的是,在瞬时遗忘发生后,只要过一段时间,我们又能回想起这些一度忘记的事情——也就是说,这些记忆确实存储在我们的大脑里,但大脑却以某种方式将它们“暂时遮蔽”了起来。
但具体是什么样的神经机制导致了这样的“遮蔽”呢?在1月21日发表于《自然》的一项研究中,来自佛罗里达州斯克里普斯研究所(Scripps Institute)的3位神经科学家揭开了这一独特机制的面纱。出人意料的是,这其中还有所谓“快乐分子”多巴胺(dopamine)的参与。
驱动遗忘的“快乐分子”
实际上,多巴胺并非简单的“快乐分子”。多巴胺是一种常见的神经调质(neuromodulator),能为大脑的不同区域传递复杂多样的全局信息。而近年,多巴胺在遗忘中的角色渐渐浮现。在一系列实验中,研究人员不仅确认了多巴胺对内源性遗忘的重要性,还找到了负责促进遗忘的多巴胺神经元,甚至连负责执行遗忘功能的多巴胺受体都得到了确认。所有的这一切,都要多亏一种常被误认为害虫的模式生物:黑腹果蝇(Drosophila melanogaster)。这种不传播人类疾病的蝇,是神经科学实验室里的常客,由于出色的繁殖能力和果蝇基因编辑的简便性,它们受到了无数实验室的青睐。
果蝇的大脑有约10万个神经元——当然,这远比不上人类大脑的860亿个神经元,但果蝇大脑的复杂程度已足以让我们利用它来探究生物记忆和遗忘。在这10万个神经元构成的大脑内有一块名为“后外侧前脑1区”(protocerebral posterolateral 1,简称PPL1)的区域,其中含有12个多巴胺神经元。这12个多巴胺神经元中,许多都向蘑菇体(mushroom body,果蝇的"学习中心")发送信息,在蘑菇体各种神经元的树突棘(dendritic spine,神经元接受信息的主要位置)上释放多巴胺,从而调控蘑菇体神经元的信号整合。
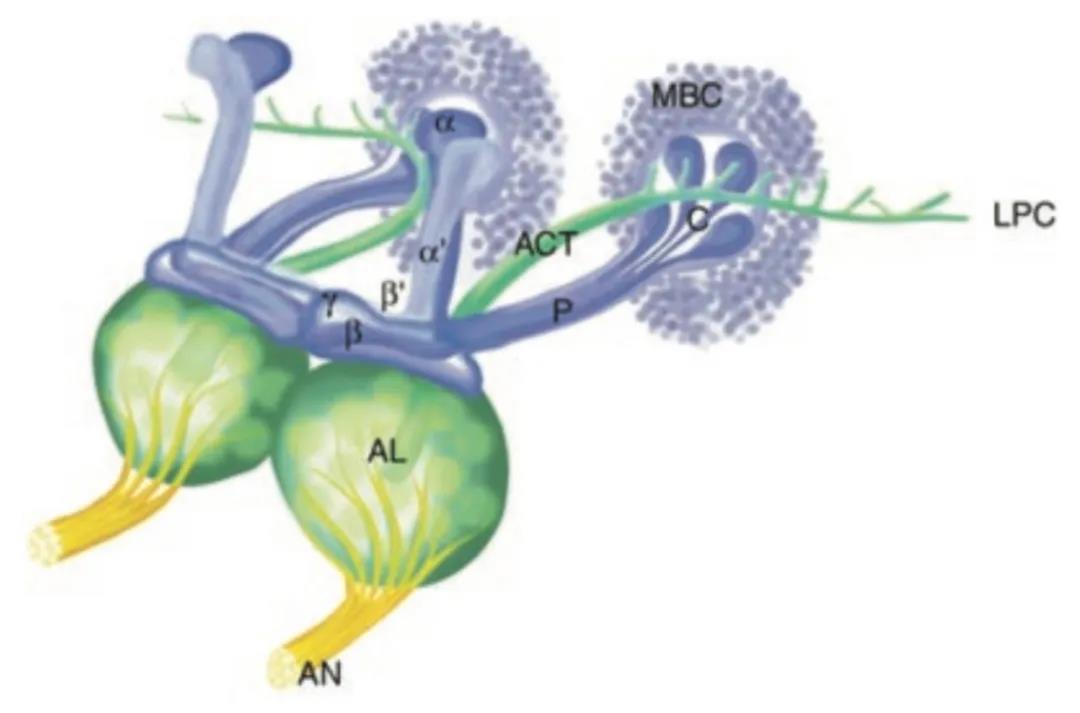
果蝇记忆系统示意图,蓝色为蘑菇体,希腊字母标注了蘑菇体的不同区域。MBC指蘑菇体细胞,ACT指触角皮质通道,LPC指侧前脑。(图片来源:McGuire, Le & Davis, Science)
神经科学家早已根据蘑菇体各种神经元的基因表达和解剖位置,将蘑菇体划分为许多大区。其中,γ2α’1蘑菇体神经元接收来自PPL1多巴胺神经元的信息,并介导内源性(长期)遗忘。过去的研究支持多巴胺介导遗忘的角色:这类有关内源性遗忘的研究,为目前这一项《自然》研究提供了宝贵的基石。
在这一项研究中,斯克里普斯研究所的罗纳德·戴维斯(Ronald Davis)与博士学生约翰·萨班达尔(John Sabandal)和博士后研究员雅各布·贝里(Jacob Berry)一起,顺着PPL1送往蘑菇体的信号,试图揭开瞬时遗忘的神秘机制。

马丁·萨班达尔和罗纳德·戴维斯教授(图片来源:斯克里普斯研究所)
果蝇:“我刚想干嘛来着?”
在这项研究中,戴维斯等人对果蝇进行了厌恶性嗅觉条件反射(aversive olfactory conditioning)训练。简单来说,果蝇会在闻到一种气味时受到轻微电击,重复训练即可使果蝇在这两者之间建立起连接——就如同巴普洛夫的狗听到铃铛后,会条件反射性地分泌唾液。与巴普洛夫的狗不同的是,经过训练的果蝇应该条件反射性地避开这种气味。怎么确认果蝇是否建立了这种连接呢?很简单,只要将训练后的果蝇放入一个T字迷宫里,在交叉路口的两边分别放上“坏气味”(“预示”了电击的气味)和“中性气味”(未与电击建立连接的气味),观察它们往哪边走就好了。结果与预想中的一样:经过训练的果蝇更“喜欢”避开"坏气味",往“中性气味”的方向走。
接下来,为了研究瞬时遗忘,戴维斯等人在果蝇选择“往左还是往右”前,短暂地对它们进行喷气、电击或蓝光刺激。他们将这些刺激统称为“干扰刺激”(interfering stimuli),目的是干扰果蝇的回忆过程,让它们在突然忘记哪个气味是“坏气味”,应该回避。实验结果毫不意外:果蝇在干扰刺激的影响之下晕头转向,完全不再回避“坏气味”。有趣的是,如果在干扰后一个小时再次测试果蝇,它们又能回忆起哪个是“坏气味”,并积极回避,选择另一条路。
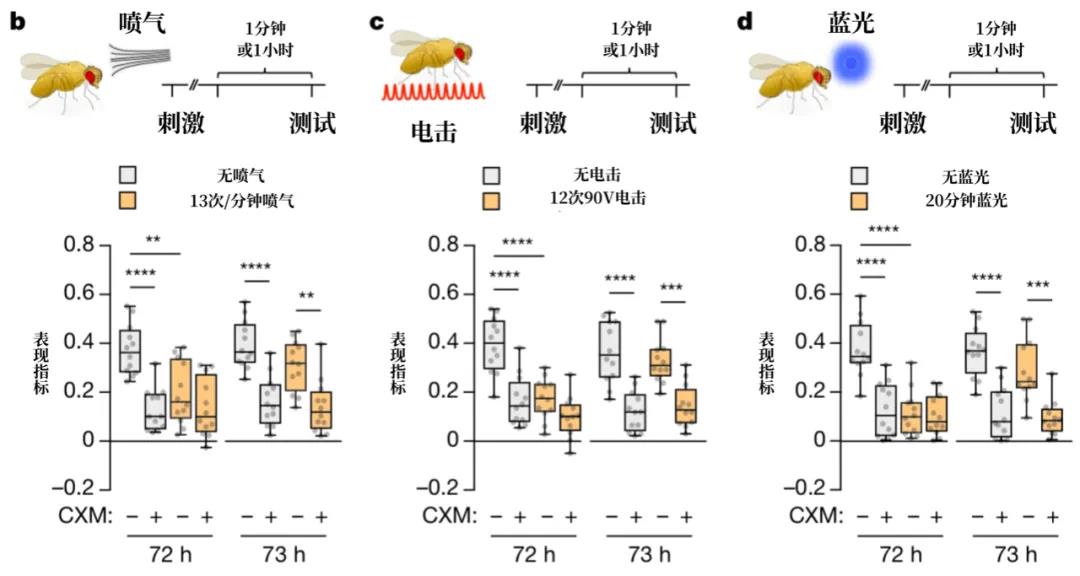
不论使用喷气、电击还是蓝光干扰,果蝇的回忆表现都会下降,即出现了瞬时遗忘。但在一小时后再次进行测试,则会发现果蝇的记忆又“回来”了。(图片来源:Sabandal, Berry & Davis, Nature,翻译制图:罗丁豪)
有了前两个实验和此前对内源性遗忘的研究,接下来的实验顺理成章。研究人员通过直接刺激PPL1中的多巴胺神经元,成功抑制了果蝇的回忆,而如果抑制这些多巴胺神经元的输出,果蝇就不会在干扰下产生瞬时遗忘。在这其中,从PPL1到蘑菇体α2α’2部分的多巴胺神经元最为重要。这以上所有证据都说明,在蘑菇体α2α’2神经元中,有一个“记忆存储器”,而多巴胺则充当“门控”(gate)的作用。多巴胺神经元的活跃,关上了回忆的大门。
从行为走到了细胞环路,戴维斯等人当然不准备就此打住。来自PPL1的多巴胺想要发挥作用,必然要刺激特定的受体;他们好奇,究竟是什么受体,什么样的细胞机制,介导了瞬时遗忘呢?
从PPL1到蘑菇体的环路上,最充沛的多巴胺受体非dDA1和DAMB莫属。此前,戴维斯实验室的研究表明dDA1受体与记忆的形成相关:果蝇的学习依赖于dDA1受体,如果没有多巴胺刺激dDA1受体,果蝇就不会记得“坏气味”。这自然留下了一个疑问:那DAMB受体呢?在使用RNA干扰技术降低α2α’2中DAMB受体的表达后,戴维斯等人发现,这些果蝇几乎完全丧失了瞬时遗忘的能力:不管怎么干扰它们,它们都能避开“坏气味”。也就是说,蘑菇体神经元的DAMB受体介导了果蝇的瞬时遗忘。
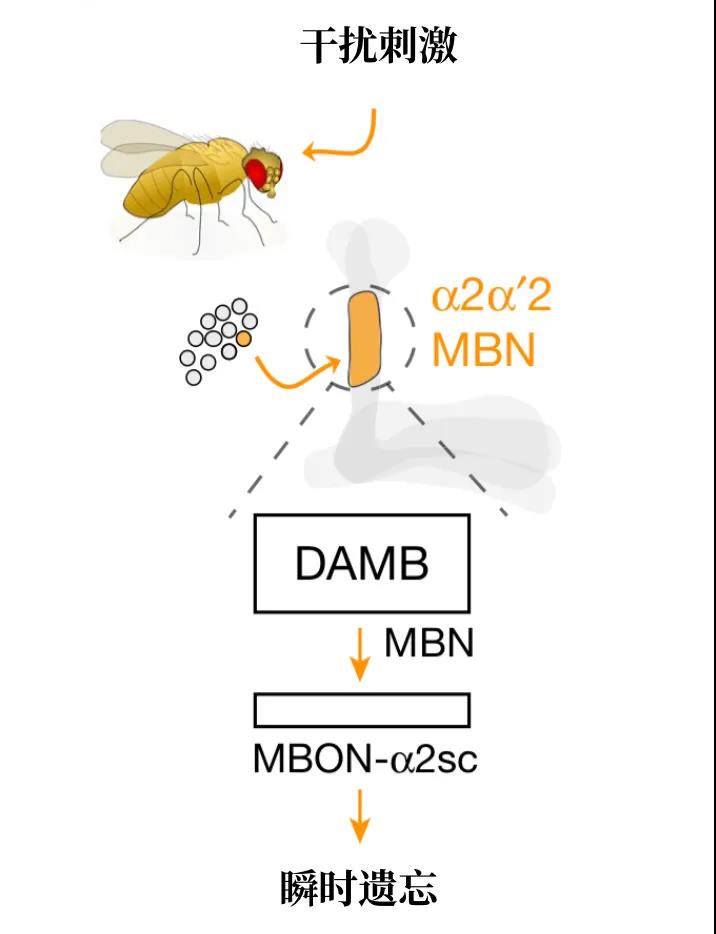
瞬时遗忘环路的示意图:干扰刺激使PPL1中的一个多巴胺神经元兴奋,这个神经元在蘑菇体α2α’2部分中释放多巴胺,刺激DAMB受体,从而介导瞬时遗忘。(图片来源:Sabandal, Berry & Davis, Nature,翻译制图:罗丁豪)
然而,在瞬时遗忘中,有一点非常重要:这种遗忘是短暂的,在数分钟到数小时后,回忆能力就会恢复。这说明瞬时遗忘(PPL1-α2α’2多巴胺神经元的活跃)并未磨灭记忆的存储,而只是简单地断开了回忆的“电路”。这也可以通过实验证实:在多巴胺神经元活跃的果蝇中,蘑菇体内的“记忆神经元”其实跟多巴胺神经元不活跃时一样活跃。也就是说,多巴胺神经元阻断了记忆的提取,但并不破坏记忆本身。
记忆容器
实际上,戴维斯实验室此前的研究已经表明,许多已被“遗忘”的记忆,实际上都以各种可塑性机制,仍然保留在细胞当中。所谓的“遗忘”,很多时候都只是“提取失败”。正在攻读博士的萨班达尔认为,遗忘只是装在“记忆容器”中的记忆“一时没办法被提取出来”。例如,在阿尔茨海默病(Alzheimer's disease)中,诺贝尔奖得主利根川进已经证明,一些“遗失”的记忆并未消失,只是被“封藏”了起来;通过合适的刺激,患有阿尔茨海默病的小鼠就能“想起”过去的记忆。当然,我们可不能无差别地记住一切事物:生命需要适应,而适应则需要关注和记住重要的事情,遗忘不那么重要的事情。因此,遗忘是生物不可或缺的技能,而瞬时遗忘可以让生物将宝贵的注意力放在应对突发事件(例如实验中的干扰刺激)上——在野外,这有时是生与死的差别。
这样的话,大脑需要高效、智能地调控记忆的形成和提取。至于多巴胺在其中的角色,戴维斯表示,我们“不清楚多巴胺究竟会对细胞做什么??我们还在瞬时遗忘研究的初级阶段。”但这项发表于《自然》的研究有力地展示了系统神经科学的优雅和美丽:从一个行为,到一个具体的神经环路,再到具体的细胞和分子机制——只有结合不同层级的研究方法,才能理解我们复杂精密的大脑。
作者:罗丁豪
编辑:顾军
责任编辑:许琦敏
来源:环球科学
声明:转载此文是出于传递更多信息之目的。若有来源标注错误或侵犯了您的合法权益,请作者持权属证明与本网联系,我们将及时更正、删除,谢谢。